
Undulatory Swimming
For a wide variety (perhaps a majority) of fishes, the dominant propulsion mechanism involves wagging or "flapping" the aft-most portion of their body, namely, their caudal or "tail" fin. Because the caudal fin is attached to the rest of the fish body, flapping of this fin is achieved by moving forward portions of the fish body (although to a lesser degree) with the net result that the fish body "undulates" as it swims. Indeed, the undulations can be described as a wave that travels down the length of the fish, reaching maximum amplitude at the end of the caudal fin.
Adapted from Gray, J. (1932) "Studies in Animal Locomotion: I. The Movement of Fish with Special Reference to the Eel," J. Exp. Biol. 10, 88-104.
Thunniform Swimming:
In many cases, the undulation is confined mostly (although not entirely) to the caudal fin. This type of undulatory swimming is called thunniform swimming for its association with tunas (genus Thunnus). Literally, "thunniform swimming" means to swim "like a tuna". Other common thunniform swimmers are whales and dolphins. Not only is action in thunniform swimming confined primarily to the caudal fin, but often (as in tunas, whales, and dolphins), the fin is crescent-shaped (lunate) like a small wing and connected to the body by only a thin section called the caudal peduncle. This unique geometry allows many thunniform swimmers to be analyzed by considering only the motion of their tail. The tail geometry and swimming motion of a dolphin are illustrated in the movie and schematic below.
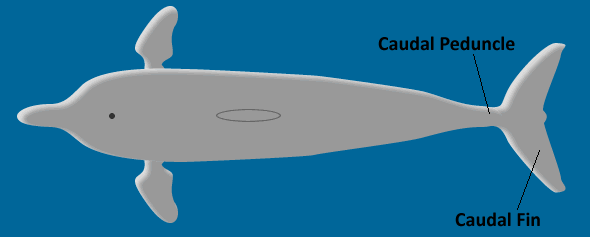
Top view of dolphin. |

Side view of dolphin. |
|
Thunniform swimming of a Bottlenose Dolphin (Tursiops truncatus). The movie shows two different dolphins swimming by in sequence. The movie was filmed at 250 frames per second (fps) with a 60 fps playback (so that the playback is slowed down by more than a factor of 4). Bottlenose dolphins live in temperate and tropical waters worldwide.
Video courtesy of Frank Fish (West Chester University) http://darwin.wcupa.edu/~biology/fish/ |
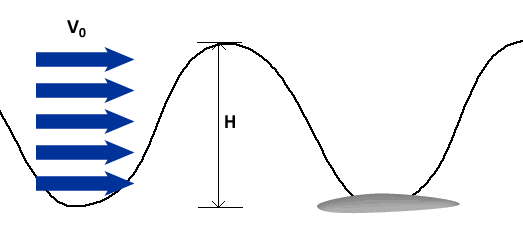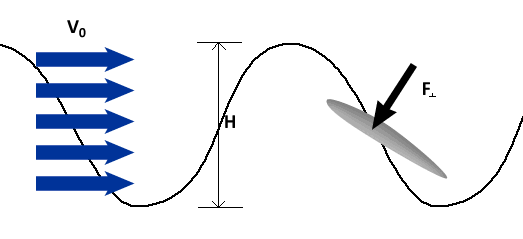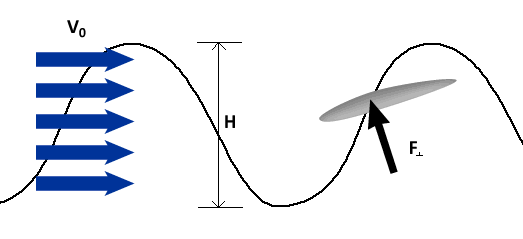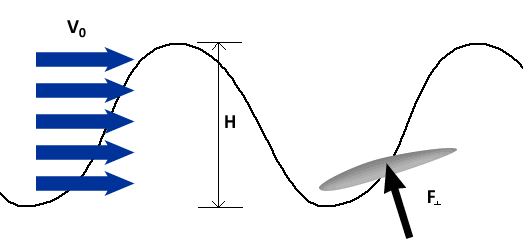
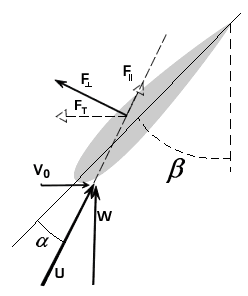
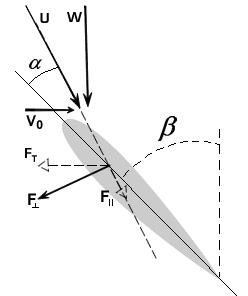
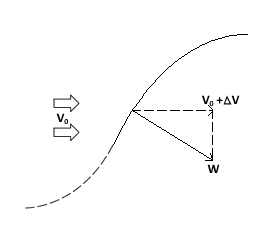
To describe the basic mechanism behind thunniform swimming, we will approximate the caudal fin as a simple symmetric wing that oscillates. From a side view, the caudal fin of a dolphin oscillates up and down as shown in the movie above and the animation below. To get a better feel for how the flow moves over the fin, it is helpful to ride along with the fin as it oscillates (click here for a review of relative motion). The flow directions from this perspective are shown in the two schematics below. The schematic on the left illustrates the flow directions for an instant where the fin is moving down at W and the fish is moving forward at V0. Then the flow velocity seen by the fin, U, is the vector sum (addition) of V0 and W. The oncoming flow U is accelerated away from the fin generating a "lift" force (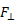 ) perpendicular to U in the same manner as a wing generates lift, as described in the section on propellers. Even though the fin is symmetric about its midline (the top and bottom of the fin have the same shape), is non-zero as long as the angle of the fin relative to U (α) is not zero. For the configuration shown, is angled forward, so its component parallel to the oncoming flow, V0, contributes thrust, FT. (Click here to review vector components.) At the bottom of the stroke, the fin must flip so it is oriented properly to create positive thrust as it moves upward on the return stroke, as shown in the schematic on the right. For this to work, the fin must be symmetric about its midline as shown. By oscillating in this manner the fin can generate positive thrust throughout the majority of the stroke.
) perpendicular to U in the same manner as a wing generates lift, as described in the section on propellers. Even though the fin is symmetric about its midline (the top and bottom of the fin have the same shape), is non-zero as long as the angle of the fin relative to U (α) is not zero. For the configuration shown, is angled forward, so its component parallel to the oncoming flow, V0, contributes thrust, FT. (Click here to review vector components.) At the bottom of the stroke, the fin must flip so it is oriented properly to create positive thrust as it moves upward on the return stroke, as shown in the schematic on the right. For this to work, the fin must be symmetric about its midline as shown. By oscillating in this manner the fin can generate positive thrust throughout the majority of the stroke.
 |
|
 Riding along with the fin as it moves down. |
 Riding along with the fin as it moves up. |
Notice the similarity between the above description and the operation of a propeller. In both cases a wing shape is used to generate a reaction force () in the same manner as an airplane wing. This force is tilted forward to create thrust by moving the wing perpendicular to the oncoming flow direction. The primary difference is that a propeller spins the wing around an axis, while a thunniform swimmer oscillates its fin back and forth. The back and forth oscillation creates sides forces that do not contribute to thrust and tend to make the animal recoil as it swims, but the side forces cancel out over one flapping cycle and the recoil tends to be minimized by other appendages on the animal, such as dorsal (top) and ventral (bottom) fins in the case of tunas.
To generate more thrust, fish can increase W, which serves to increase the magnitude of U and α. This is similar to the way propellers generate more thrust by spinning faster. For an oscillating fin, W (or more properly, the maximum W) may be increased by increasing the oscillation frequency, f, or the oscillation amplitude, H. For instance, at a fixed frequency increasing H increases W because the fin has further to go in the same amount of time. Conversely, increasing f for a fixed H increases W because the fin has less time to cover the same distance. Thus, increasing oscillation frequency and/or amplitude can increase thrust.
If W becomes too large, however, the vertical force on the fin resisting the fin motion becomes large and it requires more power to flap the fin. The increased power is analogous to the higher power required to spin a propeller faster and leads to lower propulsive efficiency. As with propeller operation, the condition for optimal efficiency occurs when α is just large enough to give the thrust required to swim, and no larger. Experiments with mechanical flapping fins show this optimum occurs at
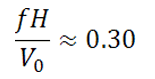
for a steady swimming. A wide variety of fishes have been observed swimming near this optimal condition at cruise, suggesting many species aim to optimize efficiency during cruise. Analysis of whale and dolphin swimming suggests they can achieve propulsive efficiencies as high as 80-90% when swimming at optimal conditions.

|
Example Problem: Fin Oscillations |
Anguilliform Swimming:
At the other end of the spectrum, undulatory swimming may involve noticeable undulation of most of the animal body, not just the tail. This is typical of eel swimming (genus Anguilla), giving it the name anguilliform swimming (swimming "like an eel"). Anguilliform swimmers often do not have a caudal fin distinct from the rest of their body (see the side view of an eel below). Instead, they move a very large fraction of their body as they swim, as illustrated in the animation and movie below.

Side of view an eel.

Top view of a swimming eel.
|
Anguilliform swimming of an American Eel (Anguilla rostrata). The eel is filmed from below (ventral view) at a frame rate of 250 frames per second (fps). The eel is swimming at about 25 cm/s or 1 body length per second. The crescent ridges at the left in the movie are shadows from ripples on the surface of the water. American eels can be found in rivers and streams along the Atlantic coast from Greenland to northern South America. Video footage courtesy of George Lauder (Harvard) |
At first glance, anguilliform swimming looks very much like wiggling a noodle or waving a ribbon. To capture this effect, we will treat anguilliform swimming as a wave that travels along the body of the fish rather than as a distinct motion of the tail. The wave starts at the head with little or no amplitude. As the wave travels back along the body, its amplitude grows. Each section of the body accelerates the fluid perpendicular to the midline of the body (pushing it "out of the way," so to speak). This is illustrated in the still image on the left below. Due to the wavy geometry of the animal body, a component of this pushing is always toward the posterior of the animal, accelerating the surrounding fluid backward to generate thrust. As the amplitude increases down the length of the animal, the fluid acceleration is increased. Of course, it is the net acceleration of the fluid above the initial speed of V0 at the head of the animal to the final value at the animal tail that governs the net thrust. Thus, we can focus mainly on the tail as illustrated on the right in the figure below, which shows the tail during a down stroke. The tail moves the fluid perpendicular to its midline at a speed W, which moves the fluid in both the horizontal and vertical directions because of the inclination of the tail. If the tail is moving quickly enough, the tail can increase the horizontal component of the fluid velocity (accelerate the flow backward), thereby generating forward thrust.
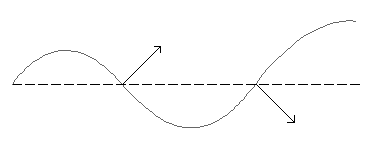 |
 |
The more quickly the tail moves (the larger W), the more the flow is accelerated and the higher the thrust. Larger W can be achieved by increasing the flapping amplitude and/or frequency. In fact, eels tend to oscillate a greater fraction of their body (increased amplitude toward the front of the body) when they try to accelerate quickly. For large thrust, it is helpful for the inclination (slope) of the tail when it flaps to be steeper. Increasing the wave amplitude along the body and having a more flexible tail will both increase the inclination of the tail and enhance thrust.
One drawback that is apparent for anguilliform propulsion is the tendency to accelerate fluid from side to side (not just backward). Under such circumstances, the animal would normally oscillate from side to side in response to the tail motion, but eels minimize this effect by oscillating a large portion of their body so that one part of the body moves in one direction while another moves in the opposite direction and the side-to-side forces tend to balance out. A more serious problem is that accelerating fluid sideways requires extra effort but does not contribute to propulsion. Equivalently, extra kinetic energy is added to the fluid without doing extra propulsive work and propulsive efficiency suffers. By oscillating just enough to get the required thrust, efficiency can be improved. Anguilliform swimmers tend to be moderately efficient, with reported propulsive efficiencies ranging from 50% to as high as 90%.
Swimming in Between:
Anguilliform and thunniform swimming represent extremes in swimming behavior. In between these two swimming modes are the carangiform (referring to swimmers from the family Carangidae, including scads and mackerels) and sub-carangiform. The range of swimming behavior is illustrated in the table below. Notice that a progressively larger fraction of the body is oscillated as you move from thunniform to anguilliform propulsion (see the swimming profiles to the left of each picture). As a general rule (with plenty of exceptions), the swimming speed increases as you move away from anguilliform swimming and most high-speed swimmers, like tuna and dolphins, are thunniform swimmers. In fact, the highest record swimming speed is for a pantropical spotted dolphin (species Stenella attenuata) swimming at 11 m/s (25 mph).

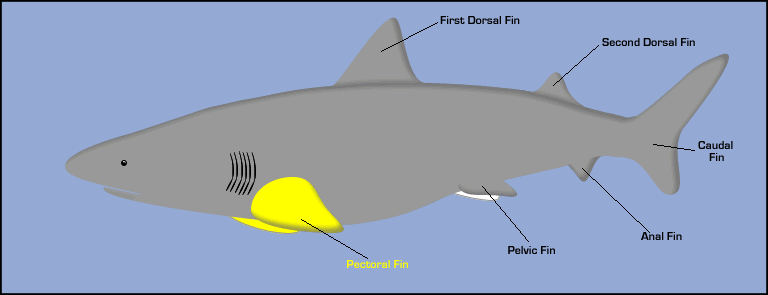
Observations based on the characteristic body motion of different swimmers highlights the basic physics involved in swimming, but sidesteps important issues related to differences in fish shape. For instance, thunniform swimmers with lunate (crescent-shaped) tails like tuna and dolphins swim using "lift" from their tails to generate thrust as described above. The tail shape is very important for optimizing propulsion in this situation. Similarly, many other fishes have very complex geometries including pectoral (side) fins, dorsal (top) fins, ventral (bottom) fins, and more. Click on the arrows in the interactive shark illustration below to learn about the functions of different fins.
 |
|
 |
|
 |
 |
For thunniform and carangiform swimmers, the dorsal and ventral fins help cancel out side-to-side motions induced by the tail flapping. They can also alter the oncoming flow so that it interacts with the tail in beneficial ways, enhancing thrust and efficiency. Pectoral fins can be used for maneuvering and stabilization, or even as the primary propulsion system at low speeds. Many species (such as bluegills and mackerels) have flexible pectoral fins that they use like oars for propulsion and maneuvering at low speeds. Clearly a detailed study of fish propulsion is a complex endeavor, but the undulatory mechanics described above covers the main physical principles utilized by a wide variety of fishes and water mammals.
